

神经现实 ✖ 哲学社 共同发布
导语 / Introduction
我们对另一个生命的道德义务至少部分取决于其精神生活的能力。我们通常通过类推来推断他者的心理状态:在某种能让我产生特定心理状态的情况下,若某个生命体和我做出的行为一致,我即可推论,该情况在TA身上产生了相同的心理状态。不幸的是,正如哲学家们早就指出的那样,这种类推是不成立的,因为行为和心理状态仅仅是偶然性相关的(contingently related)。例如,如果另一个人仅仅是在演戏,我们可能对TA的心理状态得出错误的结论。在这篇文章中我考虑了另一种形式的类推,这种类推用大脑活动代替行为,以推断对方的心理状态。根据目前大多数针对身心问题(mind-brain problem)的观点,心理状态和大脑状态是非偶然性相关的,因此,用这种新类推得出的结论可以避免行为类推的缺陷。我们用两个案例来探讨这种方法的意义:一是严重脑损伤(coma)的病人——TA们无法有意识地去做交互性的行为,二是非人动物——它们的行为方式与我们不同,且无法使用语言。由此可见,行为对于推断心理状态而言并不太有用。
关键词:心灵哲学,脑成像,持续性植物状态,动物伦理
他者心智的问题
“他者心智问题”(problem of other minds)是心灵哲学的焦点问题,它指的是人们难以得知除自己以外的其他人或物是否具有心智。以上是对这个问题的总体陈述,但它还包括各种更具体的问题。对于这些问题,我们可以通过采用不同程度的怀疑主义将其相互区分。在怀疑主义最极端的版本中,这个问题涉及到认定他者心智是否存在的困难。我们可以将其称之为他者心智的形而上学问题(metaphysical problem)。
如果假设世界上除了我们自己的心智之外还有其他的心智,那么我们就会遇到如下困难,诸如确定哪些实体具有心智,以及这些心智是什么样的。这就像我们在问“我怎么知道我的哲学教授不是一个机器人或者一个毫无心智的僵尸”时遇到的情形。怀疑主义较为温和的版本承认别人有类似于自己的精神生活,但是也指出了确定其它物种精神生活的本质困难[32]。这就是他者心智的认识论问题(epistemology problem)的诸多变种,其中涉及到从可观察的证据中推断出一种心智的存在或者其本质的困难。本文所涉及的正是他者心智的认识论问题。当然,即使是认识论问题也在很大程度上取决于关于身心关系的形而上学假设。物理证据与心理现象的推论间的相关性取决于一个人对身体和心灵间关系的看法,我将在后面再谈这个问题。
他者心智和神经伦理学有什么关联?其与伦理的关联取决于道德地位和精神生活的能力之间的关系,特别是对痛苦的感知能力。如果一个生物能感受到痛苦,那么其应该得到保护而免受痛苦。因此,我们是否能够了解、以及如何能够了解他者的精神生活是一个直接与伦理学挂钩的认识论问题。该问题与神经科学的相关性取决于神经科学的证据为我们了解生物精神生活所提供的潜在价值。在这篇文章中我将论证,在关于心-脑关系的某种形而上学的假设的背景下,神经科学的证据有别于传统上用于推断精神生活的其它证据,而且在原则上它能为我们提供更多信息。
从行为到精神状态:由类推引起的争论
他者心智的问题是心-身二元论的结果,特别体现在认为(1)身体与行为,以及(2)心理过程这两者之间没有必然联系上。笛卡尔著名的“我思故我在”表达了我们自己的精神生活确实存在的基础。但又是基于何种基础我们可推断其他人拥有思想呢?笛卡尔援引了上帝的仁慈作为相信我们关于他者心智推断的理由。如果他者心智并不存在,那么为什么上帝会给我们一个如此清晰明了的理解方式[13]?
除了神学,试图证明我们相信其他人拥有心智的做法一般基于某种类推,笛卡尔也讨论过这些,洛克[26]和其他英国经验主义者诸如约翰·密尔[29]也对此有所重视。这种类推应用了已知的自身身体与心理事件的联系,以此推断与他人可被观察到的身体事件相连的心理事件。例如图1所示,当我的脚趾被戳伤时,我会感到疼痛,并且导致我说“嗷!”当我看见乔(Joe)的脚趾被戳伤并且说“嗷!”的时候,我通过类推得出他感到疼痛。

这个类推的问题在于,它引发了另一个问题。我为什么假设在我的案例中成立的行为-心理关系在乔的案例中也成立?乔有可能只是在演戏,并没有真的觉得痛。他甚至可能是一个没有思想或者感觉的机器人。关于类似的行为-心理状态在他人身上成立的假设,即是这类类推本应该帮助我们推断的。
一个人是真的处于痛苦之中,还是在演戏,又或者只是一个机器人?这一问题可能看起来只是纯学术的探讨。毕竟常识告诉我们还不存在足够像人类的机器人能欺骗我们,而且除非有非常特殊的情况,否则也几乎没有道理去怀疑别人在演戏。然而,有两种情况让这个问题并不完全是学术性的。在这两种情况下,纵然常识阻止了我们怀疑别人是机器人或者在演戏,但对心理状态的推断似乎依然充满了不确定性。基于此,这些案例为我们呈现了关于他者心智问题的真实世界的实用版本。第一种情况是遭受了严重脑损伤的人类,他们从昏迷中醒来,对环境几乎没有行为上的反应,因此也没有通过行为来交流的能力。第二种情况是非人类动物,它们的行为反应与我们大相径庭,并且没有语言。
严重脑损伤后的认知和行为
严重脑损伤之后从昏迷中苏醒,通常符合如下模式:觉醒系统(arousal system)率先开始恢复,接着是睁眼清醒期,随后在某些情况下,或部分起伏或完全康复或恢复知觉[35]。图2展示了严重脑损伤病人可能经历的一系列状态。这些状态下的特征在表1中做了总结。
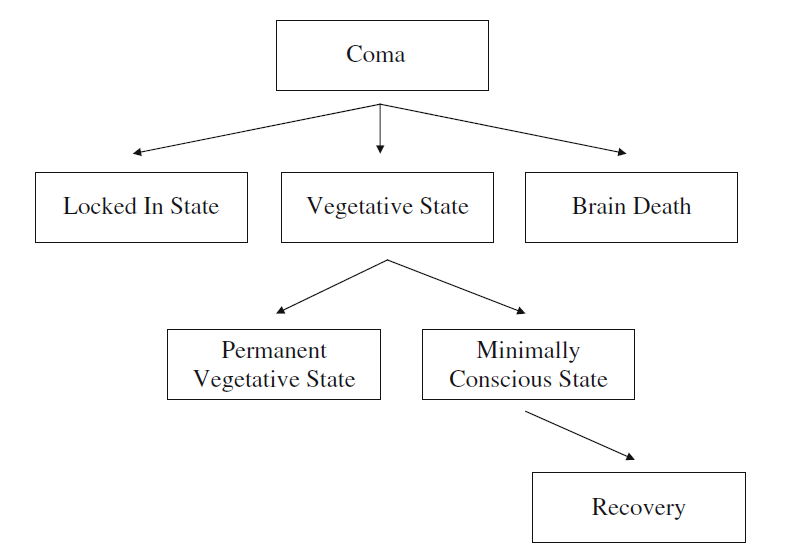
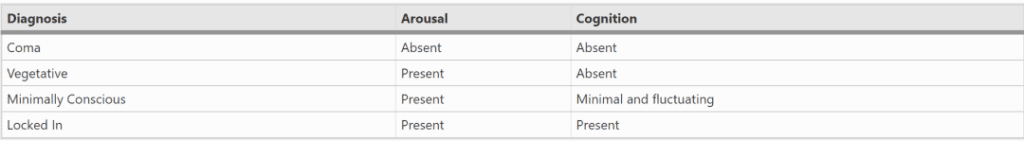
已苏醒但明显还没有意识的患者被称为“植物人状态”。他们展现出了一种令人注目的、苏醒行为与有意识行为间的解离。除了睁开眼睛之外,植物人可能会本能地挪动他们的躯干和四肢,并被观察到微笑、流泪以及发出咕哝声。他们甚至可以将眼睛和头转向周围出现可被视觉观察到的挪动或者发出声音的地方。但除了刚才提及的定向反射反应,他们对语言没有反应,他们对环境中其它方面也没有反应。
在这种情况下,他者心智的问题给病人的朋友和家人带去了沉重的压力,他们不知道他们所爱之人只是无法沟通,还是已经故去。这种不确定性带来的痛苦纷争撕裂了特丽·施亚沃(Terri Schiavo)的家庭。她的父母从女儿的行为中看到了一个能认识亲人、喜爱音乐并且想要活下去的心智。他们试图用特丽的视线变化、面部表情和其它简单可识别行为的录像带来支持他们的观点。但是她的丈夫和大多数接手这个病例的医学专家都将这些行为解释为一个无意识的人的反射性行为。
这种病人拥有精神生活的可能性带来了许多社会影响,不仅仅是使施亚沃的家人们产生分歧的维续或撤销生命支持的问题。例如,医疗机构通常只提供纯粹的监护护理,用以维持病人的身体机能,而对他们的体验没有任何关照。连续数年将一个有意识的人作为无意识的物体对待显然是不人道的。此外,在植物人患者的治疗中,不进行镇痛是很常见的[38],那么这也可能是不道德的。
精神生活对植物人状态的诊断的关键作用,以及推断精神生活的行为证据的模糊性,在最近关于这种情况的权威论述中得到了体现。该声明由持续性植物人状态多社会工作组(Multisociety Task Force on Persistent Vegetative State)做出[31]。他们将其定义为:
“一种对自我和环境完全无知觉的临床状态,伴随睡眠-苏醒周期,完全或部分保留下丘脑和脑干的自主功能(第1499页)……根据定义,处于持续性植物人状态的病人无法感知自身和所处环境。他们没有认知能力,没有知觉,也没有意识经验。”(第1501页)
他们继续陈述:
“如果断定一个人缺乏意识,而实际上他或她是有意识的,就可能出现持续性植物人状态(PVS)的错误的假阳性诊断。这种误诊可能发生在一个处于闭锁(locked-in)状态(即有意识但因严重瘫痪而无法交流)却被错误判断为无意识的病人身上。如此,从理论上说,一个看起来处于持续性植物人状态的病人可能保有意识,却没有显示出任何可以佐证的证据……在神经病学的实践中,这种情况可能性很低,所以并不影响专家们仔细建立起的临床诊断。”(第1501页)
在不贬低做出这些陈述的作者的智慧或者诚实的情况下,必须指出他们在此处提到的问题正是因为我们没有公认的方法来确定这种假阳性是否罕见。
1992年,“最小意识状态”(MCS)的诊断类别开始适用于表现出有限的、可能是间歇性的反应或交流的病人[15]。带有指示性的行为包括遵循简单指令的能力(如“眨眼”)、用语言或者手势回答“是/否”的问题、任何形式的可被理解的口语表达,或者任何与外部环境相关的有目的行为。
PVS和MCS的鉴别诊断被认为是困难的,特别是考虑到MCS中的认知的波动性。即使经过数道检查,病人间歇性、不可预测的有意识的时期也有很大可能被遗漏。因此,即使当一家医院检查病例的时候发现几乎一半的PVS诊断有误,病人确实表现出了与MCS一致的认知能力的行为证据[3],此事也并不奇怪。当然,这些假阳性的PVS的诊断并非精神状态和行为独立的结果,比如即使没有出现指示性行为,意识也可能存在。准确来说,这只是对病人行为的取样不足导致的。
相反,似乎有一些病人的认知和行为则确乎是解离了。当病人从植物人状态向最小意识状态发展时,即使他们没有表现出足以跨越意识边界的行为迹象,有些人也会展现出具有一些周期性心理活动的迹象(见[33],之后会讨论),这是有可能的。在婴儿发育过程中,获得认知能力与通过行为表现出能力之间存在类似的滞后性。这被归结为,与驱动纯粹的内部处理相比,用于驱动外部行为的内部表征在质量上有更高的要求[30]。
此外,还有另一类神经疾病患者的诊断分类,他们的认知和行为完全无关。这些人仍然能体验到对自己和对周围环境的完整的意识,但是却无法在行为上表达他们的意识。这种情况的病人被称为“闭锁”,一个令人沮丧却精准的短语,它描述了近乎瘫痪或完全瘫痪的情况,这是由于外传(传出)运动连接的中断,常见原因为中风。患者通常从昏迷中醒来后发现自己被作为植物人对待,可能在数月甚至数年内试图向医护人员和家庭成员表达自己的意识[22]。最常见的情况中,他们得以保留一定程度的自主眼动,可以用于交流,例如通过用向上的目光表示“是”来回答问题,或是用一次眼动进行单个字母选择来拼写出单词。对于另外一些病人,传出被破坏得更为彻底,自主行为已无可能[5]。
严重脑损伤患者的大脑活动
正如思考他者心智问题的哲学家一样,对于检查病人的神经学家来说,为了推断另一个生物的精神生活,行为是最明显且最自然的证据类型。然而行为从根本上来说显然是不够的,因为它与认知只是偶然的联系。此外,已知某些类型的脑损伤可以改变认知与行为的关系。
近年来,一种不同类型的证据被用于心理过程的研究,即功能性神经影像[36]。这些方法可以使神经科学家在正常人[2]和脑损伤的受试者中验证关于认知的假设[37]。这种方法最令人激动的应用之一是评估在行为上无反应或基本无反应的病人的大脑活动和大脑反应。
劳瑞斯(Laureys)、欧文(Owen)和希夫(Schiff)[19]对有关严重脑损伤之后病人脑功能的文献进行了综述。他们报告称,对静息态下全脑新陈代谢的研究表明,闭锁状态的病人大脑几乎与那些健康和清醒的人一样活跃,而处于昏迷状态、植物人状态和最小意识状态下的病人大脑活动更接近一个睡着或是被麻醉的人。正常对照组的人活动最高,随后是闭锁病人,再者是最小意识状态的病人,活动最低的是植物人。当然,全脑活动在对心理过程的研究上,相较于对(认知相关的)特定脑区活动的研究而言能揭示的东西更少,而测量静息态时大脑活动的研究比测量特定、有意义的刺激引发的活动所能揭示的东西更少。幸运的是,关于这些更具体的心-脑关联的文献数量正在增长。
与自我和环境意识有关的特定大脑区域包括前额叶和内侧顶叶皮层。这种关联是基于对正常意识和各种意识减弱如全麻、睡眠和失神发作状态下这些脑区活动的测量而得。比较本文中讨论的各种诊断门类的病人的内侧顶叶皮层在静止时的活动时,正常对照组的人活动最高,随后是闭锁病人,再者是最小意识状态的病人,活动最低的是植物人。
令人惊讶的是,拥有最小意识(MCS)的病人大脑可以保留对有意刺激的反应。例如,在Schiff及其同事的一项著名研究中[40],受试者在接受fMRI扫描的同时,向其播放亲属讲述个人相关故事的录音以及同一录音的倒放,在大致控制输入刺激的听觉特征、保留其相似性的同时,将“语言是否有意义”作为变量。与正常的对照组受试者一样,相较于倒放的无意义录音,MCS组受试者在对有意义的录音做出反应时,激活了与大脑语言相关区域的联结。作者得出结论:“一些MCS患者可能保留了广泛分布的皮层系统,尽管他们不能遵循简单的指令或者进行可靠的交流,但却有发挥认知和感觉功能的潜能。”(第514页)
相比之下,在对植物人进行的脑成像研究中,几乎没有发现任何与精神生活相关的神经传导证据。在规模最大的此类实验中,15名经仔细评估后符合持续植物性状态的病人在接受扫描时被施予疼痛刺激,与正常受试者一样,他们的中脑、丘脑和初级感觉皮层被激活。然而,与正常受试者不同之处在于,正常而言会参与疼痛刺激反应的高级皮层区域,如前扣带回(ACC)并没有被激活[21]。
对植物人的个案研究偶尔会显示出大脑保留了对有意义的刺激的反应,尽管这类研究中很少有病例在脑成像时处于明确的植物人状态。例如,一位病人的面部识别系“植物人状态上限或最小意识状态下限”[19]。迄今为止,这类文献中最引人注目的发现来自欧文及其同事对一名后来康复了的植物人患者的研究[33],但这名患者在符合植物人状态的诊断标准的同时,大脑的激活模式表明其具有语言理解能力和自主的心理想象能力。
能证明该患者保留了认知能力的一个迹象是,当向她呈现含有模棱两可词语的句子时,其大脑负责解决此类句子的脑区活动增加,这与正常人的激活区域一致。此外,当被要求执行心理想象的任务时,她的大脑活动表明她能够理解指令,并且有能力遵守指令。当被要求想象打网球时,她激活了部分运动系统;当被要求想象参观她家的每个房间时,她激活了一部分大脑的空间导航系统。此外,她的大脑激活模式和正常受试者没有区别。
总之,功能神经影像学为了解严重脑损伤患者的精神状态提供了一个新的窗口。尽管对特征明确的病人的脑成像研究还较少,但很明显,至少有一些病人虽然几乎不具备有目的的行为能力,却显示出与认知相一致的大脑激活模式。
作为精神生活的证据的行为和大脑活动
为什么当行为无法提供精神生活的证据时,大脑成像却可以?与行为相比,大脑活动是否只是一种对认知过程更加敏感的测量,但在有关精神生活的推论中却只能发挥同样的作用?还是说,它与行为测量之间有本质的差异?考虑一下大脑活动和行为扮演着类似角色的可能性。图3通过将图1中的“嗷”替换为前扣带回——大脑疼痛网络的一部分——的激活表明了这种可能性。
图3中类推的问题在于,它意味着疼痛感会导致ACC的激活,就像疼痛感导致的“嗷”一样。然而,心理状态和行为之间的关系与心理状态和大脑状态之间的关系在本质上不同。心理状态与行为间的关联是偶然性关联。一个人通过某个类似“感到疼痛”这样的短语来表达感受,这并不是一种行为,甚至不是一种行为倾向。尽管一些行为主义哲学家在数十年前就认真地探索过这些可能性,例如赖尔(Ryle)[39],但如今这并不被认为是探究心理状态术语意义的可行方法。

就了解心理状态这一目的而言,行为就像一盏指示灯。指示灯可被禁用、被切断电源,或者通过其它方式打开。它们与所指示事务间的关系取决于它们之间是否以某种方式相连接。因此,基于指示灯的推断以及类比得知,行为是不可靠的。相比之下,几乎所有当下的身心问题的学说都认为心理状态和大脑状态之间的关系是非偶然的。
在认知神经科学和当代心灵哲学中,关于心理状态和大脑状态之间关系的主流观点之一是心脑同一论:心理状态即为大脑状态。根据这一观点的一种版本——“类型同一论”(type identity)——每种类型的心理事件就是一种类型的身体事件[8, 45]。而根据另一个弱化的版本,“个例同一论”(token identity),每一个心理事件的实例都是一个身体事件的实例。最广为接受的“个例同一论”的说法基于“功能主义”(functionalism),它将某个身体状态的功能作用——在机体的输入和输出之间的中介作用——认定为相应的心理状态的决定因素[7]。功能主义自身有许多的版本,其中一些模糊了“类型同一论”和“个例同一论”之间的界限(例如[4, 24])。
有一种理论可以替代心脑同一论——基于心理状态“随附于”(supervene)大脑状态的理论,它避免了实体二元论(substance dualism),但也不认为心理状态等同于大脑状态[9, 17]。图3与随附性理论、心脑同一论均不相容。因为尽管随附性理论不认为心理和大脑状态同一,但两者之间的关系比单纯的因果关系更强。根据随附性理论,心-脑关系是非偶然相关的。用戴维森(Davidson)的话来说:“不可能存在两个事件在物理层面全然一致却在心理层面有所不同,且一件物体不可能在不改变物理层面的情况下改变其心理层面。”(第214页)
总而言之,根据当代身心问题中的所有这些形而上学立场,心理状态和大脑状态之间的关系并非偶然,就如同图1中的因果关系呈现的那样。对于类型同一论以及功能主义理论来说,例子中的前扣带回(ACC)激活与疼痛是等同的。对于随附性理论来说,如果没有疼痛,ACC的激活就不可能存在。因此,将大脑活动与心理状态的推论绘制成图4那样会更有道理。这个图的要点在于,无论你对乔大脑中的ACC激活有多少确信的成分,你都可以肯定他处于痛苦中。简言之,由大脑活动的推论得出的论证对那些干扰行为推论的其它解释是免疫的。
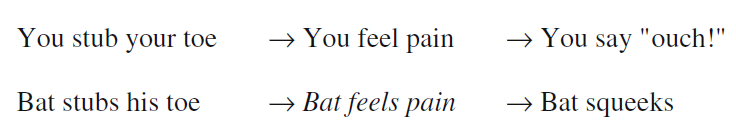
他者、非人动物和心智的问题
与之前讨论的严重脑损伤病人一样,非人动物的交流能力有限,这种限制令我们无法使用常用的方法来了解它们的心理状态[1, 11]。尽管今天很少有人会同意笛卡尔关于动物完全缺乏心理状态的结论,但我们大多数人却对动物精神生活的本质与程度感到不太确定。一方面,我们许多人将某些动物拟人化,尤其对我们的宠物,我们根据人类在相同情况下可能的想法,将复杂的思想和期待映射到它们身上。另一方面,动物的精神生活常常被认为是假设的,和我们人类的精神生活无法媲美,甚至根本不存在。否则我们如何接受用粘鼠板捕捉啮齿动物以及煮熟龙虾当晚餐的行为?
如图5所示,非人动物为我们提供了他者心智问题的另一个版本,由于人类和动物的行为方式不同,有问题的类推显得更有问题了。动物无法说话,甚至不能以与人类相近的非语言方式表示痛苦。例如,有的动物根本不会发声,而且有的动物在害怕的时候有可能僵住而非颤栗。
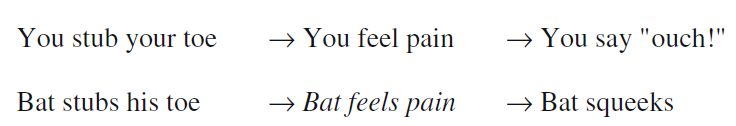
神经科学的方法能否为我们探索其它生物的精神生活提供指引?在某种程度上来说,它已经提供了,然而根据目前的分析,它还能提供更多指引。伦理学家曾将生理学数据用于处理动物痛苦的问题,特别是有关人类与动物痛觉系统的相似性上。例如,Singer([42]第12-13页)详尽地引用了一位痛觉研究者的著作,大意为疼痛处理是一种较低层次的大脑功能,人类在这方面和其它动物差异不大。这种对生理学数据的使用与现今的使用相比,有两方面不同。
首先,根据目前的分析,生理学数据不只是关于生物精神生活的又一个证据来源,它需要和行为证据一并被权衡,从而在这方面可能更有价值。或许正如上一节所述,由于生理学数据与心理状态间的非偶然关系,它们可以发挥一种在性质上不同且更具确定性的作用。就前述图表而言,这就是图3和图4的区别。
第二个区别来自于认知神经科学相对更前沿的能力,即将大脑过程拆解为有意义的心理和伦理类别。在目前的案例中,它揭示了所谓“单纯疼痛”和痛苦之间的神经基础差异。疼痛具有很多不同的方面,其中一个与伦理学特别相关的方面是痛苦的心理质量[10, 12, 16]。有的疼痛体验主要来自于身体,有的则来自于心理痛苦。道金斯(Dawkins)用“既不愉快又强烈”形容后者,与“痛苦”这个词语相称。根据参与疼痛的脑区的不同,疼痛可以划分出生理和心理成分——与疼痛状态相对应的神经状态似乎遵循了这一重要区别。
对动物和人类的研究表明,在对疼痛刺激的反应中,一个分布甚广的大脑区域联结变得活跃,包括丘脑和躯体感觉皮层区域、也包括那些进一步远离感觉输入的区域,如脑岛和前扣带回(ACC)。当疼痛的物理强度变化时,例如在脑成像实验中,让人类受试者触摸一个致人疼痛的可变温表面,整个联结的活动水平会发生变化[6]。利用人类可以报告自己心理状态的能力(以及原则上第一人称研究的可能性,即心理状态的内省),我们能分别将疼痛的心理和生理维度作为变量,并绘制出各维度的对应大脑状态。例如,吗啡被公认可减少心理部分的疼痛。被施用吗啡的病人通常报告说他们仍能感觉到“生理”疼痛,但这种疼痛已经不那么困扰他们了。参与催眠暗示治疗疼痛的病人也报告了相同的情况。这两种干预措施主要在前扣带回区域有神经方面的效用[18, 25, 34]。当没有遭受疼痛的人对遭受疼痛的人强烈共情时,即使他们没有遭受疼痛,他们的ACC也会被激活[44]。这些发现表明ACC的激活反映了痛苦(suffering)而非“单纯”疼痛(pain)。
施利弗(Shriver)[41]指出哺乳动物拥有ACC,因此在神经层面上具备心理和身体疼痛的能力。按照施利弗的说法我们可以用动物来代替图4中的乔。然而,由于只有拥有相似大脑的生物之间的大脑状态才可能相似,我们必须修改图6中的图表,以规定人类和动物的ACC只适用于各自的情况。

这就引出了一个问题:在一个物种的大脑中确立的心-脑关系能否推广到其它物种。这个问题的背后是一个更根本的问题,即大脑状态的变化程度和类型如何对应心理状态的变化程度和类型,这个问题即使在同一个物种内也会出现,因为没有哪两个大脑是完全相同的。原则上来说,人可以控制人类大脑中的变量(包括自己的大脑),以系统性地改变令每个人大脑有所不同的生物物理特征,从而探究大脑状态的哪些方面负责决定心理状态。当然这在实践中完全不可能。
我们最多可以假设心理状态的相似性会随着大脑状态的相似性的下降而下降,在不知道这些相似性与大脑状态的哪一方面相关,也不知道其中一个状态的下降对另一个状态的影响几何的情况下。埃德尔曼(Edelman)、巴尔斯(Baars)和塞斯(Seth)提供了一个例子[14],试图确定不同物种(包括非哺乳动物)大脑结构的功能相似性。施利弗[41]试图通过引用ACC在大鼠和人类疼痛历程中发挥类似作用的证据,以解决从人类向非人类泛化的问题(尽管这种证据公认是基于行为之上的,而目前向脑成像证据的靠拢是为了取代基于行为的证据):拉格莱兹(LaGraize)及其同事[20]比较了健康大鼠和ACC损伤大鼠的行为,迫使大鼠在处于黑暗中和避免被电击脚之间进行选择(大鼠一般喜欢待在黑暗中,但黑暗处有电极)。所有的大鼠在受到电击时都收回了被电的脚,这表明它们保留了对疼痛的感知。然而脑损伤的大鼠更愿意为了留在装有实验装置的黑暗区域而经受电击。就像服用了吗啡的病人一样,它们似乎对疼痛不那么敏感。这意味着大鼠的ACC发挥的作用与人类的ACC相当。
我是在暗示神经科学可以告诉我们当一只蝙蝠是什么样子吗?是,也不是。当内格尔(Nagel)[32]提出这个问题时,他选择蝙蝠作为非人类动物的代表,因为蝙蝠用回声定位来感知世界,这是人类缺乏的。即使有神经科学的帮助,对我们而言,用我们不具备的感官来感知世界的模样仍然存在问题,因为在人脑中没有蝙蝠进行回声定位的神经系统的同源组织。然而,鉴于我们和蝙蝠都有相同的一般性的疼痛生理学,包括ACC,因此我们可以知道作为一只蝙蝠的某些方面。具体来说,我们可以推断,和一个脚趾受伤且注射了吗啡的人类相比,一只脚趾受伤的蝙蝠更像一个脚趾受伤且没有止痛药的人(一只脚趾受伤的蝙蝠更像一个脚趾受伤且没有止痛药的人类,而不是一个脚趾受伤但注射了吗啡的人类)。
迄今为止,动物心智的问题在神经伦理学中还不够受重视。其中一个原因或许是因为神经伦理学还是一个年轻的学科,尚未涉及到最终将构成该学科的所有领域。另一个原因可能是,和动物伦理学相关的个人和政治争议使学者们不愿意接近这个话题。鉴于动物伦理学在现实世界中的重要性,以及神经科学证据在这一学科发展中的特殊作用,动物神经伦理学的研究看起来有很大的前景。
假设和结论
“神经科学可以揭示关于严重脑损伤病人和非人动物伦理的相关信息。”上述想法是基于一些假设的。本文尚未研究的一个假设是,我们对一个生命的伦理义务至少应部分取决于其精神生活。尽管这个假设几乎是毋庸置疑的,但关于精神生活的一些具体方面有哪些具体的伦理含义还有待商榷。也许最为重要的下一步澄清关涉到精神生活中那些使我们有义务遏制痛苦的方面,以及那些使我们有义务保护生命的方面。
本文重点讨论了另一个生物是否具备相对简单的心理状态的问题——那些具备一定意识体验和情感价值的心理状态。与感知自己和构想自己的生活、并明确倾向于继续生活的心理能力相比,这种心理能力的伦理意义更为有限[23, 43]。迄今为止的神经科学证据仅限于呈现病人和非人动物有体验前一种心理状态的能力,因此相关的伦理意义局限于遏制痛苦而非保护生命。然而,这并不是使用神经数据这一方法本身的局限性:如果有适当的研究计划,我们完全可以确定与“自我的概念和继续生活的愿望”相对应的神经系统及其状态。这种知识将对临终决策的许多方面产生影响,甚至可能使我们负有不可杀害某些动物的义务。
另一个值得明确讨论的假设涉及认知加工过程与意识之间的关系。认知神经科学家在成像实验中将认知加工过程和大脑激活联系起来。目前,这是一个重要的假设,因为我们在伦理上的关注点在于有意识的精神生活,特别是有意识的痛苦,而不是无意识的信息处理。根据前文有关心脑关系的主流观点,特定神经过程的类型或实例,与某些心理状态(包括有意识的心理状态)等同,或必然与之相关。因此,问题在于,凭借经验科学的手段确定哪些大脑状态对应哪些有意识的心理状态。这不是个小问题,但原则上是可以解决的。事实上,如果人们愿意接受其他正常人对有意识的体验的报告作为证据,我们在实践中就可以解决这个问题。(不愿意接受别人关于有意识体验的报告的怀疑论者不妨对自己的大脑做脑成像。这可以用来验证特定的发现,但不可能作为验证所有认知神经科学知识的手段。)
最后一个假设涉及认知神经科学的准确性和完整性。为了探讨神经科学证据作为他者心智问题的解决方案在原则上的前景和局限性,我在写作本文时假设了我们已经得知与特定心理状态相关的大脑状态。很不幸的是,这并非事实。尽管认知神经科学在过去几十年中取得了长足的进步,但我们目前的知识状态还远远谈不上“完整”。对于很多心理状态,包括痛苦,我们对相关脑区有很好的工作假说(working hypotheses),但未来的研究无疑会对其中一些假说进行修改。此外我们对这些脑区执行相关心理状态的具体机制知之甚少。脑成像研究中观察到的“激活”与单个细胞水平测量到的神经活动密切相关,但并不能完美地对应神经元行为的具体方面,如动作电位[27]。此外,对大脑活动的任何单一的测量,无论是对单个细胞还是细胞集合,电传导还是化学传导,都会遗漏神经元功能的潜在重要特征。用我们目前的方法测量出的激活有可能无法判断出相关的神经元活动,并且在某些情况下可能产生误导。了解大脑成像涉及区域的神经元进行的计算,包括它们与其它区域神经元的相互作用,对评估跨物种同源性的尝试而言尤为重要。
参考文献
1. Allen, C. 2004. Animal pain. Nous 38: 617–643.
2. Aguirre, G.K. 2005. Functional imaging in cognitive neuroscience I: Basic principles. In Patient-Based Approaches to Cognitive Neuroscience, eds. M.J. Farah, and T.E. Feinberg 2nd Cambridge: MIT Press.
3. Andrews, K., L. Murphy, C. Munday, and C. Littlewood. 1996. Misdiagnosis of the vegetative state: Retrospective study in a rehabilitation unit. British Medical Journal 313: 13–16.
4. Armstrong, D.M. 1968. A Materialistic Theory of the Mind. London: RKP.
5. Bauer, G., F. Gerstenbrand, and E. Rumpl. 1979. Varieties of the locked-in syndrome. Journal of Neurology 221: 77–91.
6. Becerra, L.R., H.C. Breiter, R. Wise, R.G. Gonzalez, and D. Borsook. 2001. Reward circuitry activation by noxious
thermal stimuli. Neuron 32: 927–946.
7. Block, N., and J. Fodor. 1972. What psychological states are not. Philosophical Review 81: 159–181.
8. Churchland, P.M. 1988. Matter and Consciousness. 2ndCambridge: MIT Press.
9. Davidson, D. 1970. Mental events. Reprinted in essays on actions and events. Oxford: Clarendon Press.
10. Dawkins, M.S. 1985. The scientific basis for assessing suffering in animals. In In Defense of Animals, ed. P.
Singer, 27–40. New York: Basil Blackwell.
11. DeGrazia, D. 1996 Taking Animals Seriously. Mental Life and Moral Status. Cambridge University Press.
12. Dennett, D.C. 1996. Kinds of Minds: Toward an Understanding of Consciousness. New York: Basic Books.
13. Descartes, R. 1968. Discourse on Method and the Meditations (trans. F. E. Sutcliffe). Penguin.
14. Edelman, D.B., B.J. Baars, and A.K. Seth. 2005. Identifying hallmarks of consciousness in non-mammalian species. Consciousness and Cognition 14: 169–187.
15. Giacino, J.T. et al. 2002. The minimally conscious state: definition and diagnostic criteria. Neurology 58: 349–353.
16. Hardcastle, V.G. 1999. The Myth of Pain. Cambridge, Massachusetts: MIT Press.
17. Kim, J. 2005. Physicalism, or Something Near Enough. Princeton, NJ: Princeton University Press.
18. Kupers, R., M.E. Faymonville, and S. Laureys. 2005. The cognitive modulation of pain: hypnosis- and placebo-induced analgesia. Progress in Brain Research 150: 251–269.
19. Laureys, S., Owen, A., and Schiff, N. 2004. Brain function in coma, vegetative state, and related disorders. The Lancet Neurology 3: 537–546.
20. LaGraize, S.C., C.J. LaBuda, M.A. Rutledge, R.L. Jackson, and P.N. Fuchs. 2004. Differential effect of anterior cingulate cortex lesion on mechanical hyperalgesia and escape/avoidance behavior in an animal model of neuropathic pain. Experimental Neurology 188: 139–148.
21. Laureys, S., M.E. Faymonville, P. Peigneux, P. Damas, B. Lambermont, G. Del Fiore, C. Degueldre, J. Aerts, A. Luxen, G. Franck, M. Lamy, G. Moonen, and P. Maquet.
2002. Cortical processing of noxious somatosensory stimuli in the persistent vegetative state. Neuroimage 17: 732–741.
22. Laureys, S., F. Pellas, P. Van Eeckhout, S. Ghorbel, C. Schnakers, F. Perrin, J. Berre, M.E. Faymonville, K.H. Pantke, F. Damas, M. Lamy, G. Moonen, and S. Goldman. 2005. The locked-in syndrome: what is it like to be conscious but paralyzed and voiceless? Progress in Brain Research 150: 495–511.
23. Levy, N. 2008. Cognition and Consciousness. in press.
24. Lewis, David. 1980. Mad pain and Martian pain. In Readings in the Philosophy of Psychology, Vol. I., ed. N.
Block, 216–222. Harvard University Press.
25. Lidstone, S.C., and A.J. Stoessl. 2007. Understanding the placebo effect: contributions from neuroimaging. Molecular Imaging & Biology 9: 176–185.
26. Locke, J. 1959. Essay Concerning Human Understanding, ed. A.C. Fraser. Dover.
27. Logothetis, N.K., and B.A. Wandell. 2004. Interpreting the BOLD signal. Annual Review of Physiology 66: 735–769.
28. Menon, D.K., A.M. Owen, E.J.Williams et al. 1998. Cortical processing in persistent vegetative state. Lancet 352: 200.
29. Mill, J.S. 1979. The collected works of John Stuart Mill, volume IX—An examination of William Hamilton’s philosophy and of the principal philosophical questions discussed in his writings, ed. John M. Robson, Introduction
by Alan Ryan (Toronto: University of Toronto Press, London: Routledge and Kegan Paul, 1979). Chapter: CHAPTER XII: The psychological theory of the belief in matter, How far applicable to mind. Accessed from http://oll.libertyfund.org/title/240/40871 on 2007-12-13
30. Munakata, Y. 2001. Graded representations in behavioral dissociations. Trends in Cognitive Sciences 5: 309–315.
31. Multi-Society Task Force on PVS. 1994. Medical aspects of the persistent vegetative state (1). New England Journal of Medicine 33021: 1499–1508.
32. Nagel, T. 1974. What is it like to be a bat? Philosophical Review 83: 435–450.
33. Owen, A.M., M.R. Coleman, M. Boly, M.H. Davis, S. Laureys, and J.D. Pickard. 2006. Detecting awareness in
the vegetative state. Science 313: 1402.
34. Petrovic, P., E. Kalso, K.M. Petersson, and M. Ingvar. 2002. Placebo and opioid analgesia—imaging a shared
neuronal network. Science 295: 1737–1740.
35. Plum, F., and J.B. Posner. 1980. The diagnosis of stupor and coma. 3rdPhiladelphia: FA Davis Co.
36. Posner, M.I., and M. Raichle. 1997. Images of mind. New York: WH Freeman.
37. Price, C.J., and K.J. Friston. 2005. Functional imaging in cognitive neuroscience II: Imaging patients. In Patientbased approaches to cognitive neuroscience, eds. M.J. Farah, and T.E. Feinberg 2nd Cambridge: MIT Press.
38. Rifkinson-Mann, S. 2003. Legal consequences and ethical dilemmas of pain perception in persistent vegetative states. Journal of Health Law 364: 523–548.
39. Ryle, G. 1949. The concept of mind. Chicago: University of Chicago Press.
40. Schiff, N.D., D. Rodriguez-Moreno, A. Kamal, K.H.S. Kim, J.T. Giacino, F. Plum, and J. Hirsch. 2005. fMRI reveals large-scale network of activation in minimally conscious patients. Neurology 64: 514–523.
41. Shriver, A. 2006. Minding mammals. Philosophical Psychology 19: 443–442.
42. Singer, P. 1990. Animal Liberation, revised edition. New York: Random House.
43. Singer, P. 1999. Practical ethics, 2nd ed. Cambridge University Press.
44. Singer, T., B. Seymour, J. O’Doherty, H. Kaube, R.J. Dolan, and C.D. Frith. 2004. Empathy for pain involves the
affective but not sensory components of pain. Science 303: 1157–1162.
45. Smart, J.J.C. 1959. Sensations and Brain Processes. Philosophical Review 68: 141–156. M.J. Farah
作者:Martha J. Farah | 翻译:Phosphas
审校:Zen、杨银烛 | 封面:Moon皓玥
编辑:杨银烛、星原 | 排版:光影
原文:https://repository.upenn.edu/neuroethics_pubs/35/














 沪公网安备31011202020814号
沪公网安备31011202020814号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
评论