
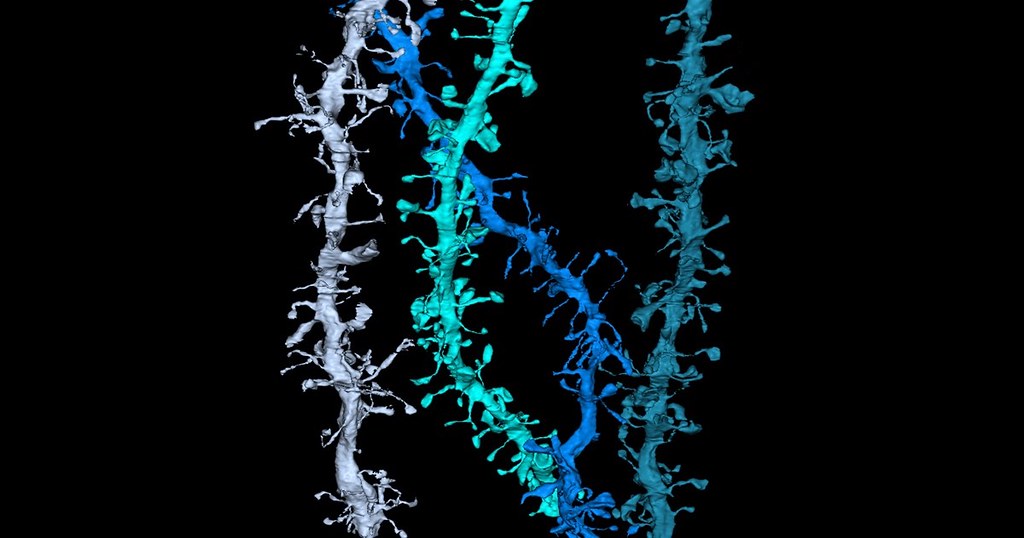
最近,博利厄·拉罗什(Beaulieu-Laroche) 等人和吉东(Gidon)等人都报告了他们首次在人类新皮质神经元的树突上进行了膜片钳记录。这些研究表明,人类的树突表现出电敏感性,在树突上可以观察到反向传播的动作电位以及快速的树突钙离子发放。这一新的前沿研究提示了一种可能性:树突计算的生物物理学性质存在跨物种差异。
从树突功能的跨物种差异性窥探人类之谜
单个神经元是如何运作的?对于这一问题,我们的很多理解都来自于对模型系统(model systems)的研究。对模型系统的选取,往往会考虑到实验的便利。其中,在大鼠、猫、水蛭、龙虾、枪乌贼和蛞蝓的神经元上进行的实验,帮助我们回答了一些有关神经元运作的分子和细胞层面上的基本问题。许多问题在所有的神经系统中都具有普遍性——这些问题可以通过研究模型系统来解答。
但模型系统解答不了某些人类独有的问题。其中,就包括一些对于临床转化具有明显重要性的问题,例如:人类神经递质受体有哪些药理学性质?对这些性质进行直接测量,可能对于研发新药物至关重要。然而,研究人类神经元的实验,除了和临床相关之外,更重要的是,它们组成了我们长期以来为了理解人类的认知所做出的努力中的一个必要部分。我们卓越的认知能力可能来源于在细胞和网络两个层面的适应性,并且在某些情况下,可能会依赖于特定的,会造成功能限制的生物物理性质。
树突占据了神经元表面的绝大部分,并且它们表现出很多有可能会带来上述功能限制的性质。大多数突触都处在树突上,因此,树突的一些被动和主动性质决定了神经元对突触输入的整合,进而影响神经元动作电位的输出。树突的特殊之处,可以给神经元的输入-输出功能带来两种不同变化:一种是变得线性而简单,另一种是变得完全非线性且复杂[1]。因此,人类树突的任何特性都可能会影响到神经元和神经环路所执行的计算,并且可能会带来人类认知上的特殊化。此外,哺乳动物的大脑中含有数量庞大的神经元和树突,这也就意味着:即使是在树突功能上相对小的量上的差异,也可能会转化成人类大脑显著的计算优势。
人类的神经元和树突与其他动物(例如猫和大鼠)神经元的树突有一个明显区别:它们更大。在这种解剖结构上的差异,本身就可以改变树突的信号处理功能[2,3],但这种差异是否也伴随着功能性质上的差异?早期在神经外科手术(例如为了治疗癫痫或脑肿瘤的手术)中,在切除的皮质组织上进行的胞内记录表明,人类神经元在很大程度上与其他哺乳动物神经元在功能上是相似的[4]。然而,较新的膜片钳研究,让人类和其他哺乳动物的神经元之间一些有趣的差异初现端倪。例如,单独的一个动作电位可以引发一连串的局部网络活动,这些网络活动在人类的皮层组织中持续的时间,比在啮齿目动物的皮层上要久[5]。并且,与啮齿目动物的神经元相比,人类的2/3层神经元具有更低的膜比电容值(specific membrane capacitance)[6]。然而,人类的树突是否也表现出功能差异,这一点尚不清楚。
对人类树突的膜片钳记录揭示跨物种差异性
近日,有两个研究团队获得了重大突破:第一个团队在脑片上对人类的锥体神经元树突进行了膜片钳记录,揭示了人类树突的功能特性。博利厄·拉罗什(Beaulieu Laroche)的团队报告,人类与大鼠皮质第5层的锥体神经元相比,树突发生兴奋的过程相似,但人类的树突更不易兴奋,并且也更长。这一特点改变了胞体和树突的输入-输出性质。具体来说,人类第5层神经元的尖端树突具有更强的电绝缘性,对于这些离胞体较远的树突区域来说,这一性质可能会增强它们的计算独立性,潜在地提供了更加丰富的计算指令库[1]。
与拉罗什在第5层神经元树突上的研究不同,吉东(Gidon)等人[8]发现人类的树突比啮齿目动物的2/3层的树突更容易兴奋:研究者向这些神经元的树突中注入电流,结果产生了重复的快速的树突钙发放串(spike trains),这些发放串独立于胞体的动作电位。像这样的树突发放,在随时进行的计算以及突触可塑性的触发中具有重要作用[1]。吉东等人发现的钙发放,与啮齿目动物小脑神经元的发放相似[9],也和啮齿目动物2/3层神经元树突的发放相似[10],但又表现出了一些关键的不同之处。有趣的是,研究表明,这些发放事件对于注入电流的强度有矛盾的双相敏感性(biphasic sensitivity)——更高程度的刺激会带来反应幅值的降低。在此之前的研究表明,树突可以执行逻辑运算[11]。作者提出了一个简化的模型来说明,从原则上来讲,对输入强度表现出双相敏感性的树突,可以执行异或(exclusive-or,XOR)运算。在运算中,树突的发放事件会有选择性地增强较低程度的输入,而不会增强更高程度的输入。
一些有待深入研究的新问题
这些具有开拓性的研究表明,人类皮质树突具有电敏感性,并且与其他物种的树突具有诸多相似性。然而,这些研究同时也强调了一些有趣的跨物种差异,启发了很多新的问题。第一,人类和啮齿目动物树突之间的功能差异究竟是质上的还是量上的?换句话说,这些差异会表现为生物物理上相区分的树突事件,还是说它们仅仅是一个连续体上的两个不同的点?要回答这个问题,研究者需要采用完全相同的方案,进行仔细的、并行的跨物种对比研究。而人类组织样本的稀有性使得研究者很难去控制它们之间的差异:不同的疾病状态,大脑区域,细胞类型,药理学史,甚至病人的年龄和认知状况都必须考虑在内。要一般化人类皮层上的不同种类的神经元就已经很困难了:最近的研究中存在一些值得注意的明显的不一致,例如在膜比电容值(单位膜面积的电容值)以及突触兴奋性的问题上的矛盾,这些问题还需要被仔细地核实和被更深入地探索。
*译者注:
1.盖·埃亚勒等人[6]在人类颞叶皮层2/3层的锥体神经元上测得的膜比电容值约为0.5mF/cm2,这一数值仅为人们普遍认为的生物膜的膜比电容值(约1mF/cm2)的一半;与之相对的,博利厄·拉罗什等人[7]在人类外侧前颞叶第五层的神经元上记录到的膜比电容值约为0.9mF/cm2,与大鼠无异。
2.博利厄·拉罗什等人[7]报告说人类皮层第5层的锥体神经元比啮齿目动物相应神经元的树突兴奋性要弱;而艾伯特·吉东等人[8]则报告说人类皮层2/3层的锥体神经元的树突兴奋性比啮齿目动物相应神经元的树突兴奋性更强。
第二,这些所被观察到的“特殊化”的差异特性究竟是独特存在于个别物种的,还是说,这些生理机制广泛存在于动物王国,而我们所捕捉到的仅仅是一些例子?要理解像这样更宏大的组织法则,需要在很多物种之间进行仔细的对比。第三,我们所观察到的功能差异的生物物理学起源是什么,它们又是如何与人类和非人神经元之间存在的分子、解剖层面的差异相联系的?回答这一问题需要在许多不同层面进行一连串的系统性的实验,包括对转录和药理学的研究(Box 1)。最后,在完整的脑内,人类突触的计算性质是怎样被利用的?回答这一问题最终会需要对人类树突进行在体记录(而这面临着相当大的实践上和伦理上的挑战),进一步作为补充,还需要在大型网络中对具有真实的树突性质的神经元进行仿真模拟。回答这些问题可以让我们准确地分辨出,我们的树突和大脑在哪些方面区别于其他物种,它们在哪些方面又与其他物种一样服从普遍的准则。这两方面的研究结果都会为理解人类认知带来贡献。
跨物种树突性质对比清单
对人和非人类的树突进行系统性的对比是非常有价值的,其价值不仅体现在疾病的诊断和治疗方面,还体现在帮助人们理解人类树突的信息处理,在何种程度上可能是独一无二的,又在何种程度上反映了动物王国的普适法则。在某些情况下在人类身上做实验存在困难(例如,一些在体测量),在非人灵长目(尤其是猕猴和狨猴)上进行记录同样也会提供有用的信息。
在这里,我们把要进行严格的跨物种对比所需要的信息总结起来,列出了一个“清单”:
- 对树突结构的形态学特性描述
- 人类和非人神经元的转录组
- 树突上的通道和受体的分布情况
- 树突的位置和mRNA的运输
- 树突兴奋性的功能特性
- 被动的电张性质
- 反向传播的动作电位的扩散
- 树突发放的普遍性和类型
- 突出输入的模式整合
- 在树突上的钙和第二信使的信号通路
- 突触可塑性的机制
- 在行为过程中对树突信号的在体测量。如果这样的实验已经可以在人类身上在体实施,研究者应该用它们来探索树突兴奋性是怎么服务于与行为相关的群体活动的,理想情况下这些实验最好可以在复杂的认知任务过程中进行。
最后,以上的信息必须要与神经元以及网络的数学模型整合起来,这样才能更严格地探究实验结果中所得到的计算准则。还可以将这些结果植入到一个定量的框架中,这样做同样也能让我们检验所观察到的人和非人树突的差异究竟是属于同一个连续体还是代表着相互区别的特征;同时借此我们也可以窥探到,生物是经历了怎样的演化过程而获得一些特定属性的。
参考文献
[1] Häusser, M. and Mel, B. (2003) Dendrites: bug or feature? Curr. Opin. Neurobiol. 13, 372–383
[2] Bekkers, J.M. and Stevens, C.F. (1990) Two different ways evolution makes neurons larger. Prog. Brain Res. 83, 37–45
[3] Vetter, P. et al. (2001) Propagation of action potentials in dendrites depends on dendritic morphology. J. Neurophysiol. 85, 926–937
[4] Foehring, R.C. et al. (1991) Correlation of physiologically and morphologically identified neuronal types in human association cortex in vitro. J. Neurophysiol. 66, 1825–1837
[5] Molnár, G. et al. (2008) Complex events initiated by individual spikes in the human cerebral cortex. PLoS Biol. 6, e222
[6] Eyal, G. et al. (2016) Unique membrane properties and enhanced signal processing in human neocortical neurons. eLife 5, e16553
[7] Beaulieu-Laroche, L. et al. (2018) Enhanced dendritic compartmentalization in human cortical neurons. Cell 175, 643–651
[8] Gidon, A. et al. (2020) Dendritic action potentials and computation in human layer 2/3 cortical neurons. Science 367, 83–87
[9] Llinás, R. and Sugimori, M. (1980) Electrophysiological properties of in vitro Purkinje cell dendrites in mammalian cerebellar slices. J. Physiol. 305, 197–213
[10] Larkum, M.E. et al. (2007) Dendritic spikes in apical dendrites of neocortical layer 2/3 pyramidal neurons. J. Neurosci. 27, 8999–9008
[11] Koch, C. et al. (1983) Nonlinear interactions in a dendritic tree: localization, timing, and role in information processing. Proc. Natl. Acad. Sci. U. S. A. 80, 2799–2802
译者:Orange Soda
审校:阿莫東森
原文:https://doi.org/10.1016/j.tics.2020.03.002












 沪公网安备31011202020814号
沪公网安备31011202020814号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
评论